Why aging is editable now
Aging is not a single switch that flips off after youth. It is a layered systems failure that shows up as DNA damage, epigenetic drift, mitochondrial wear, chronic inflammation, and cells that refuse to retire when their shift is over. Synthetic biology treats that mess like software bugs. It senses what is wrong, computes a response, and actuates a fix using programmable parts. The field has matured from speculative gene tinkering to a design discipline with standards, safety layers, and factory lines for building living circuitry.
This guide translates the state of play into plain language. It spans sensors that read biological age, gene and RNA circuits that respond to those signals, living and nonliving delivery vehicles, and early clinical paths where the first wins are most likely.
The new toolkit at a glance
Sensing. Aging clocks based on DNA methylation, single‑cell RNA and chromatin profiles, and proteomic patterns that reveal which cells are old, stressed, or senescent.
Computation. Synthetic gene and RNA circuits that integrate those signals with logic rules. Examples include inducible switches, feedback controllers that keep dosage in range, and kill‑switches that shut systems down if they drift.
Actuation. Programmed effectors that rejuvenate cells, remove the harmful ones, or bolster failing subsystems. These include epigenetic reprogramming payloads, senolytics, designer immune cells, mitochondria editors, and engineered exosomes or probiotics.
Delivery. Viral vectors such as AAV for long‑lived expression, lipid nanoparticles for transient or tissue‑targeted RNA delivery, and living carriers like engineered microbes or vesicles. Control layers add timers, sensors, and off‑ramps for safety.
What aging looks like from a circuit designer’s perspective
A synthetic biologist breaks aging into detectable states and controllable transitions.
- State 1: Recoverable dysfunction. Mitochondria, proteostasis, or metabolism is off, but the cell can still be nudged back into a youthful program.
- State 2: Senescence. The cell is permanently growth‑arrested and secretes inflammatory factors. Here the safe play is removal.
- State 3: Maladaptive remodeling. Tissues like heart, lung, or brain have rewired around damage. They need coordinated, multi‑node interventions.
Circuits sense these states, then choose among three high‑level actions: reprogram, remove, or replace.
Reprogram: dialing back cellular age without erasing identity
Partial epigenetic reprogramming uses subsets of developmental factors to reset gene expression and DNA methylation patterns while keeping the original cell type intact. In multiple mouse models, inducible delivery of OSK factors has rejuvenated tissues and improved frailty measures late in life. In the eye, targeted expression has restored vision and regeneration in models of optic nerve injury and age‑related degeneration. Several groups have now reproduced long‑term functional recovery in retinal ganglion cells and are advancing ophthalmic indications because the anatomy allows local dosing and careful monitoring.
Design lessons for engineers:
- Use tight, drug‑inducible control. Overexpression or the wrong tissue can tip cells toward cancer or loss of identity.
- Dose in pulses, not floods. Short, spaced inductions steer cells toward youthful programs and away from dedifferentiation.
- Add safety rails. Self‑deleting editors, reversible RNA switches, and time‑limited vectors reduce risk.
Where this likely lands first: diseases with clear, local delivery windows such as nonarteritic anterior ischemic optic neuropathy and glaucoma. Next wave: cartilage and intervertebral disc repair where cells are accessible and contained.
Remove: precision senolytics that target the right cells at the right time
Senescent cells accumulate with age, drive chronic inflammation, and impair regeneration. Clearing the harmful subsets improves tissue function in many models. Small‑molecule senolytics were the first proof, but synthetic biology adds precision.
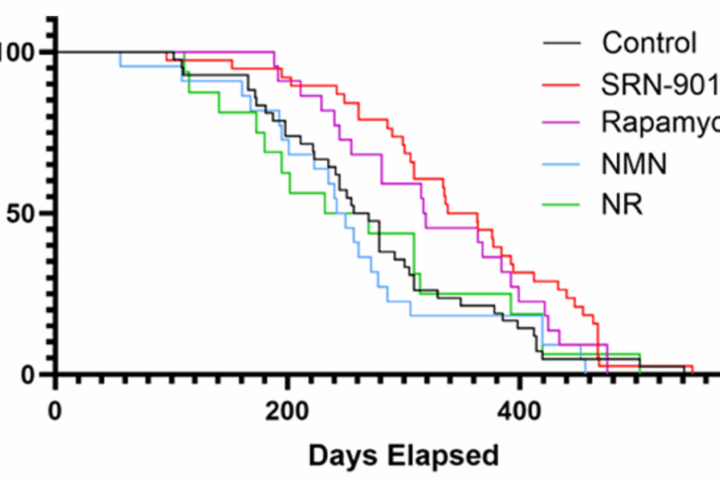
Promoter‑driven gene therapies. DNA constructs under the control of senescence‑associated promoters like p16 or p53 can express apoptotic effectors only in target cells. In aged mice, combinations of these constructs have reduced senescence burden across tissues, improved frailty scores, and extended remaining lifespan. The attraction is obvious: redosable, systemically delivered senolysis that ignores healthy cells.
Senolytic CAR‑T cells. T cells engineered to recognize senescence‑linked surface markers have ablated senescent hepatocytes and fibrotic cells in mouse models and reversed pathology. Next‑gen versions gate activation using logic rules, which raises specificity and lowers collateral damage.
Engineered exosomes and vesicles. Vesicles decorated with senescent‑cell homing peptides or antibodies can carry siRNAs, microRNAs, or small‑molecule payloads to remodel the tissue environment without permanent genetic change.
Vaccination strategies. Virus-like particle (VLP) and nanovaccine platforms that elicit antibodies against senescence‑enriched surface proteins are emerging as a low‑cost, scalable complement. These approaches are attractive where chronic dosing is acceptable and where humoral immunity does most of the work.
Engineering notes:
- Senescence is heterogeneous. Combine promoters, receptors, and secreted cues to avoid missing key subtypes.
- Add “deadman” switches to kill or quench the therapy if off‑target activation appears.
- Track outcomes with standardized senescence panels, not single markers.
Replace and repair: mitochondria, proteostasis, and tissue microenvironments
Mitochondrial genome editing. Base editors that operate in mitochondria can correct point mutations or model them with high precision. For aging, the nearer‑term use is to restore respiratory chain balance in tissues like muscle and brain. Delivery remains the main bottleneck, so expect local or ex vivo applications first.
Proteostasis controllers. Synthetic circuits that upregulate autophagy, chaperones, or the unfolded protein response can be tuned to rescue stress without triggering apoptosis. Yeast lifespan experiments that rewired aging circuits into an oscillator show the value of feedback control, not brute force.
Microbiome engineering. Next‑generation probiotics and engineered bacterial vesicles can produce short‑chain fatty acids, bile acid modulators, and neurotransmitter precursors on schedule. Think of them as living pharmacies that reinforce host resilience and immune tone.
RNA is a circuit language now
Programmable mRNAs can compute inside cells. They carry logic that turns therapeutic protein expression on only when a target signature is present and turn it off if the message lands in the wrong tissue. This is a natural fit for aging, where context matters. You can imagine an mRNA therapy that reads a senescence microRNA code, produces a rejuvenation factor only in those cells, then self‑silences.
Advantages:
- Transient by default, so safer for first‑in‑human studies.
- Scalable manufacturing that already meets global vaccine standards.
- Easy to retarget as biomarkers improve.
Delivery and control: the hidden half of the problem
Therapeutics fail more from imprecise control than from weak biology. Modern designs layer multiple safety and control features.
- Inducible expression. Small‑molecule controlled switches or ribozyme on‑switches let clinicians dial exposure up or down.
- Feedback controllers. Circuits keep expression within a therapeutic band, which prevents both underdosing and toxicity.
- Self‑limiting editors. Cas9 and base editors that remove themselves after editing lower long‑term risk.
- Kill‑switches. Genetic fuses terminate therapy if stress signatures or off‑target patterns appear.
- Targeted carriers. AAV serotypes, receptor‑targeted lipid nanoparticles, or cell‑homing exosomes put payloads where they matter.
Safety, ethics, and the regulatory path
Aging itself is not a formal disease category in current drug frameworks, which is why most near‑term programs lead with organ‑specific diseases. The big risks are dedifferentiation and tumorigenesis from over‑reprogramming, insertional mutagenesis from uncontrolled integration, off‑target editing, immune reactions to vectors, and long‑lived expression when you only want a pulse. The countermeasures are getting sharper: tighter inducible systems, self‑deleting editors, logic‑gated cell therapies, and validated senescence reporting standards.
Ethically, access and misuse matter. Many of these platforms can be repurposed. Guardrails, transparency, and post‑market surveillance should be built in from day one.
Where the first wins are most likely
- Eye diseases with local delivery. Nonarteritic anterior ischemic optic neuropathy and glaucoma are natural on‑ramps for partial reprogramming because the retina and optic nerve can be targeted precisely and monitored.
- Fibrosis and osteoarthritis. Senolytics, reprogramming pulses, and matrix‑modulating vesicles can be layered. Success criteria are functional, not just histology.
- Cardiometabolic disease. Vascular and adipose senescence is tractable with promoter‑driven gene therapies, logic‑gated CAR‑Ts, or vaccines against senescence‑enriched surface markers.
- Pulmonary and kidney disease. Senescence contributes to chronic inflammation and remodeling in COPD and CKD. Targeted senolytics plus tissue repair programs can shift trajectories.
How to read the next two years like an insider
Evidence standards. Favor programs that report multi‑omic aging clocks, functional outcomes, and safety telemetry, not a single marker. Look for blinded analyses and pre‑registered protocols.
Control layers. Prefer payloads with drug‑inducible expression, self‑deleting editors, or kill‑switches. These are hallmarks of serious translational intent.
Delivery realism. Be wary of whole‑body claims without a clear delivery story. Local or compartmentalized indications are more credible first steps.
Combination logic. The most compelling designs stack reprogramming to reset, senolysis to clean up, and metabolism or proteostasis support to stabilize the gains.
A brief field notebook of concrete examples
- Gene oscillators that extend lifespan. In yeast, rewiring the aging network into a negative feedback oscillator avoided runaway failure states and raised lifespan dramatically. The punchline is not the yeast, it is the control theory.
- Inducible OSK gene therapy late in life. Very old mice given pulsatile OSK expression showed large gains in remaining lifespan and mobility, with organ‑level rejuvenation markers. In the eye, targeted OSK restored vision and regenerative capacity in injury and age‑related models across multiple labs.
- Senolytic gene therapies. Aged mice given systemic, promoter‑gated senolytic constructs cleared harmful cells, improved frailty metrics, and even reduced spontaneous tumor incidence, suggesting immune tone improves when the worst cells are gone.
- Senolytic CAR‑Ts. T cells aimed at senescence‑linked surface proteins rapidly eliminated pathological cells and reversed disease signatures in liver and lung models. Gated designs promise even tighter control.
- Mitochondrial base editors. Editors that never enter the nucleus can make precise C to T or A to G changes in mitochondrial DNA. In vivo studies have shown tissue‑specific editing, which opens a path to treat bioenergetic failure.
- Programmable mRNA therapies. Logic‑gated mRNAs turn therapeutic protein production on only inside cells with a specific signature and self‑silence elsewhere. Oncology is leading, but the same logic applies to aging tissues.
- Engineered exosomes and probiotics. Vesicles that home to senescent cells deliver microRNAs that ease fibrosis and inflammation in preclinical models. Probiotics engineered to make short‑chain fatty acids or bile acid modulators on cue improve metabolic resilience in animal studies.
What success will feel like
It will not be a single fountain. It will be a series of quiet clinical wins where people keep their sight, breathe easier, climb stairs without pain, and avoid hospitalizations. The platforms will stack. A short course of reprogramming will pair with a senolytic gene therapy and a probiotic that stabilizes immune tone. The result will be fewer bad cells, better energy management, and tissues that behave like they used to.
If biology is the hardware, synthetic biology is the operating system update. We are finally learning how to ship patches that stick, without bricking the device.
References
- López-Otín, C., Blasco, M. A., Partridge, L., Serrano, M., & Kroemer, G. (2023). Hallmarks of aging: An expanding universe. Cell, 186(1), 4–27. https://doi.org/10.1016/j.cell.2022.11.001 PubMedunifal-mg.edu.br
- Ajoolabady, A., Praticò, D., & Ren, J. (2025). Hallmarks of cellular senescence: Biology, mechanisms, regulations. Experimental & Molecular Medicine. https://doi.org/10.1038/s12276-025-01480-7
- Zhou, Z., et al. (2023). Engineering longevity—Design of a synthetic gene oscillator to slow cellular aging. Science, 380(6649), eadd7631. https://doi.org/10.1126/science.add7631
- University of California–San Diego. (2023, April 29). Scientists slow aging by engineering longevity in cells. ScienceDaily. https://www.sciencedaily.com/releases/2023/04/230427173454.htm
- Amor, C., Feucht, J., Leibold, J., Ho, Y.-J., Zhu, C., … Lowe, S. W. (2020). Senolytic CAR T cells reverse senescence-associated pathologies. Nature, 583(7814), 127–132. https://doi.org/10.1038/s41586-020-2403-9
- Amor, C., Fernández-Maestre, I., Chowdhury, S., Ho, Y.-J., Nadella, S., … Lowe, S. W. (2024). Prophylactic and long-lasting efficacy of senolytic CAR T cells against age-related metabolic dysfunction. Nature Aging, 4(3), 336–349. https://doi.org/10.1038/s43587-023-00560-5
- Silva-Pinheiro, P., Kannan, S., Gonzalez-Garay, M. L., et al. (2022). In vivo mitochondrial base editing via adeno-associated viral delivery of DdCBE. Nature Communications, 13, 4247. https://www.nature.com/articles/s41467-022-32428-1
- Ogrodnik, M., Adams, P. D., Acosta, J. C., Demaria, M., & Grillari, J., et al. (2024). Guidelines for minimal information on cellular senescence experimentation in vivo (MICSE). Cell, 187(16), 4150–4175. https://doi.org/10.1016/j.cell.2024.05.059
- Tao, W., Yu, Z., & Han, J.-D. J. (2024). Single-cell senescence identification reveals senescence heterogeneity, trajectory, and modulators. Cell Metabolism, 36(7), 1126–1143.e5. https://doi.org/10.1016/j.cmet.2024.05.004
- Mullin, E. (2024, January 22). Scientists will test a cancer-hunting mRNA treatment. WIRED. https://www.wired.com/story/cancer-hunting-mrna-programming-treatment-test/
- Wyss Institute at Harvard. (2025). Application Area: Healthy Aging (incl. Ichor: Reversing Aging). https://wyss.harvard.edu/application-area/healthy-aging/
- Financial Times. (2025, March). Sam Altman-backed Retro Biosciences to raise $1bn for project to extend human life. FT.com. (Paywalled) Financial Times
- Zhou, Z., et al. (2023). Engineering longevity—Design of a synthetic gene oscillator to slow cellular aging (author’s PDF). Science. https://haolab.ucsd.edu/zhou_science_2023.pdf haolab.ucsd.edu